











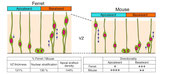


三次元培養下にとらえた,神経前駆細胞の分裂の様子
左の2枚組パネルは,脳原基のapical面付近においてtangentialな光学切片を得られるよう共焦点顕微鏡観察を行って得られたライブ像(脳室の側からapical面より数ミクロン奥の世界を眺めた)である.細胞膜が可視化できるような蛍光色素を用いて全細胞(それぞれに疑似カラー)の輪郭を描出している(Dev. Dyn. 236, 3061-3070, 2007; Dev. Growth Differ. 50, S105-S112, 2008).中央(疑似カラーなし)の丸い大きな部分がM期(分裂中)の細胞体であるが,それが分裂して2つの娘細胞が誕生する様子が分かる.細胞分裂に伴って,周囲の細胞突起(疑似カラー)が「そよぐ」ように四方に押し出される(ちなみに「押し出し」「そよぎ」の様子は,右の組み写真パネル中にも発見できる).
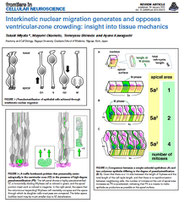
右の長い「組みパネル」は,左のパネルとは直交する断面を見るように行われたライブ観察(cross-sectionalなスライス作製がなされた)例である.単一の神経前駆細胞(左端のタイムポイント)を蛍光標識し,観察を始めた.神経前駆細胞はapical面(画面下)からbasal面(画面上)まで伸びた細長い形態をしている.神経前駆細胞はapical面で分裂し(2番目のタイムポイント),2つの娘細胞を生み出した(Dev. Growth Differ. 45, 219-229, 2003).娘細胞の片方が,「親」細胞からbasal突起を丸ごと相続した.どちらの娘細胞も,誕生地であるapical面からいったん深部(basla側)へと核・細胞体を移動させ,やがて再びapical面に戻り分裂した.こうした核の「行き戻り」運動は「interkinetic nuclear migration (INM)」と称される.
INMは,1897年にSchaperが意識し,1935年にSauerが唱え,1959年にトリチウムチミジンを用いた「パルス・チェイス法」によって証明された.1995年にChenn & McConnellによってフェレットのスライス培養下に神経前駆細胞の INM が初めてライブでとらえられた(Cell 82, 631-641).2001年にNoctorらがラットの(Nature 409, 714-720),Miyataらがマウスの(Neuron 31, 724-741)スライス培養を行い,INMを行った前駆細胞がapical面で「長いまま(basal突起を保有したまま)分裂する」と示した.細胞産生に対して主体的に貢献するのが,こうした「長い」細胞(従来はニューロン移動のガイド役としてのみ知られ「放射状グリア」と称されてきた細胞)であると認められるようになったのはこの年からである.
「見えた」ことにもとづいて,新たな問いが 湧き出す
さて,INMのメカニズム(核がどうやって動くか)については最近,研究が進んできた.微小管,アクトミオシンによる制御が考えられている(総説:Dev. Growth Differ. 50, S105-S112, 2008; Neuron 67, 906-914, 2010; EMBO J. 30, 1676-1677, 2011; J. Mol. Neurosci. 46, 516-526, 2012; Cell. Mol. Life Sci. [in press, Kosodo]; 当研究室でも関連する報告をしてきた:Genes Cells 12, 983-996, 2007 [東北大 大隅研との研究]; Neurosci. Res. 63, 294-301, 2009). また,apical面に向かう核の「押し」に依存して受動的に周囲の核がapical面から移動するとの機構(EMBO J. 30, 1690-1704, 2011 [Kosodo et al.]; Development 138, 5003-5013, 2011 [Noden team])も考えられている.
上右パネル中で,basal突起を相続した娘細胞と相続しなかった娘細胞では,INMの様子に違いがあるように見える(basal突起相続細胞のINM軌跡を赤く,非相続細胞のINM軌跡を青で示す)ので,INMに対してbasal突起が貢献していると予想されるのだが,これまで直接実験的に問われた事はなかった(他の状況を変えずにbasal突起を無くすという操作を加える事は技術的に困難であるので).当ラボでは,現在,この点について新しい成果を得つつある(2012年 3月「Neocortical Organization」,4月 日英合同 発生生物・細胞生物学会,5月 日本発生生物・細胞生物合同学会,9月 日本神経科学会などで発表).
「押され(押し)」の観察(左の組みパネル)は,機械的・力学的な関係性が細胞たちの営みに何らかの意味を持つ事を暗示する.さらに,実は,組織の中には「引っ張られ」もある.ニューロンの軸索が引っ張られながら伸びるとの報告がある(J. Neurosci. 24, 7978-7983, 2005)が,「長い神経前駆細胞」(放射状グリア)にも引っ張り力が作用していることが分かっており(Curr. Biol. 17, 146-151, 2007),それが前駆細胞の細胞産生の活動性に対して何か影響を与えるのだろうかと,不思議に思う.
さまざまな建築物におけると同様に,脳という組織づくりにも「圧縮力と張力の組み合せ」が潜んでいると想像される(2012年 1月 定量生物学の会 年会,5月 日本発生生物・細胞生物合同学会で発表)のだが,そのような視点での探究は,まだ方法論の模索の段階であり,今後,学際的な取り組みが必要だと考えている.
さて,現在のところ,INMの意義(動く事が何に役立つか,逆に動かぬとどう困るのか)について,いくつかのアイディア(網膜の形成過程について唱えられる「未分化性の維持」機能--------- Cell 134, 1055-1065, 2008-------など)が提示されてはいるが,詳しくはほとんど分かっておらず,これからの研究が求められる.当ラボでの現在進行中のプロジェクト(2012年 3月「Neocortical Organization」,4月 日英合同 発生生物・細胞生物学会,5月 日本発生生物・細胞生物合同学会,9月 日本神経科学会などで発表)は,この点に対してもひとつの答えを示すことができるものと考えている.
ところで,apical面で誕生して途中までは(basal向きの局面は)INMを遂行するがやがてapical面との結合を断って「旅立つ」娘細胞もいる.そうした「旅立ち娘細胞」には「ニューロンづくり専門的な(lineage restrictedな)前駆細胞」(Development 131, 3133-3145, 2004) やニューロン(Neurosci. Res. 57, 326-329, 2007)が属する.この「旅立ち」の機構は,まだ分かっていない.
ひとつ分かっているのが,「旅立ち」は,ほとんどの場合は「誕生」の瞬間に起きるのではない,ということである.つまり,apical面で誕生したほとんどの娘細胞は,誕生後しばらく(数時間)の間,apical面につながっている.この状態で,一部の娘細胞に Neurogeninなど,ニューロン分化の方向付けにかかわる転写因子が発現し始めることが分かっている(Mol. Cell. Neurosci. 40, 225-233, 2009 [胎生中期の大脳皮質原基を対象とした研究]).
いったんNeurogeninがしっかりと発現し始めると,「旅立ち直前」あるいは「旅立ち中」の細胞たちは,周囲細胞に対してデルタ(Notch のリガンド)を提示する(Development 135, 3113-3124, 2008 [川口,CDB松崎研にて]; Neuron 58, 519-531, 2008 [韓国グループとの共同研究]).本来はこのようにして起動・進行するはずのNotchシグナル経路が遮断されると,集団としての「バイナリー」な運命選択が破綻する(未分化な前駆細胞の維持が叶わず,ニューロンづくりの方向に偏る)ことが分かっている.
ではそのNeurogeninの発現はどうやって始まるのか?というところが,まだ分かっていない(Curr. Opin. Neurobiol. 20, 22-28, 2010).ただ,上記の「旅立ち直前」あるいは「旅立ち中」の細胞たちは,デルタの発現を介するなどして,彼らにとって少し(数時間)だけ「後輩」であるところの「apical面で誕生したばかりの娘細胞たち」にとって重要な「環境」になっている.「後輩」細胞(apical面で誕生したばかりの娘細胞たち)のうちの一部のみが「Neurogenin発現」に至りそれ以外のものが「Neurogenin非発現」との意を決するまでには,こうした「環境」が深く関与していると考えられる.
「細胞間相互作用」に向き合う
このように,娘細胞の運命選択には,「細胞間相互作用」の関与が強く意識されている.しかし,現時点では,「集団レベルの運命選択」と,「個々の分裂,個々の娘細胞の運命選択」とを結び付ける事が,まだ,決して,できてはいない.
「50個の前駆細胞が分裂して,100個の娘細胞が誕生するとき,50個が分化し50個が未分化である」というポピュレーションレベルで「バイナリーな(二方向性の)運命選択」がなされるとして,では,「いま私が見ている『この』神経前駆細胞の分裂で生じたクローン(A)の2つの娘細胞たちの運命決定と,そのすぐそばで起きた『別の』前駆細胞の分裂によって生じたクローン(B)の中での運命選択が,いずれも『片方の娘細胞が分化,片方が未分化』というバイナリーなものになっているか」ときちんと問われた事は,これまでほとんどなかった.
ひとつの可能性として考えるなら,「クローンAでは2つの娘細胞の両方が分化,クローンBでは2つの娘細胞がともに未分化」という状況であっても「集団としてはきれいにバイナリー」ということが成立し得る.仮にそのような事が起きるとするなら,それはどうやって果たされているのだろう.
よく「非対称分裂」を考えるモデルとして,「分裂する個々の細胞において例えば地球の北半球と南半球に分けるような運命決定因子の相続の有無」が論じられる.そのパターンが50ケースの分裂で50回再現されるなら,その機構にはきわめて決定的な意味があると見なすことができる.しかし,私たちはそのパターンを安心して「50個の分裂細胞」にコピー&ペーストしてよいという保証を持っているのだろうか.
当ラボがスライス培養を用いて胎生中期のマウス大脳皮質原基のapical面で分裂した前駆細胞から誕生した「2細胞クローン」39ケースを解析した結果,「分化・分化」クローン(注:娘細胞の「分化」の判定は Neurogenin2の抗体染色による検出にもとづく)が21%,「未分化・未分化」クローンが26%存在し,「未分化・分化」といういわゆる典型的な「非対称」なクローンは54%にとどまった(Mol. Cell. Neurosci. 40, 225-233, 2009). また,その後,レトロウイルスを用いた in vivo の解析でも類似の結果(「未分化・未分化」および「分化・分化」2細胞クローンの存在)を得て,2011年 日本神経科学会で発表を行なった.
さらに,「非対称」クローンも,どちらの娘細胞が「分化」なのかという目で見れば,決して均一とは見なせないと考える.よく注目点として取り上げられる「basal突起の相続・非相続」が「娘細胞の未分化・分化」と対応するかという問題(総説:Cell 146, 18-36, 2011など)では,「相関」の度合いが例えば80%などと高かったとして,では,「残りの20%(「basal突起の相続」と「未分化」が対応しない,「真逆」のケース)」をどう説明すれば良いのかと,素直に悩まざるを得ない.仮に「突起相続」にまつわる「未分化促進機構」が働いているにしても,それ以外の機構も同時に(「真逆」パターンを生じさせるように)働いているのではないかと思わずにはいられない.当ラボでは,そういう「悩ましい」ケースの実在に,かなりの頻度で遭遇している(Mol. Cell. Neurosci. 40, 225-233, 2009;2011年 日本神経科学会で発表)ので,「何かひとつ」ですっきりとシロクロがつくというようにして娘細胞の運命の決定が行なわれるとは,なかなか考えにくい.
つまり,我々は,単純な「コピー&ペースト」はできないだろう,と思っている.
上述の「デルターNotch」の問題について,我々は,「あるポピュレーションにおいてデルタ提示が乏しいと,そのポピュレーションから生じる娘集団で未分化性の維持ができなくなる」ことをかなり前から知っている.ちなみに,ごく最近(4月12日)出版されたGuoチームによるゼブラフィッシュ胚を用いた研究では「単一の前駆細胞に由来する4細胞クローン」というごく小さな「ポピュレーション」にまで分解能が高められている(Neuron 74, 65-78, 2012).
しかし,ある「単一の娘細胞」に関して,「先輩」からのデルタ提示(したがって,自身でのその受け取り)が多い場合に「未分化(Neurogenin非発現)」,そうでない場合に「分化」の道を選択しているのか,と逐一の納得ができているのかと言うと,実はそのような分解能での理解は,まだない.
娘細胞にとっての「場」「環境」のありようについて,我々の理解は依然「抽象的・文学的」である.「細胞間相互作用」と呼び捨てることで「無名化」し,ひとまず満足せざるを得ない段階にいるように,私には思える.どうにかして,これに個別性を持たせることができるような観察を行い,「相互作用」のあれこれ,すべてを見尽くし,集団の掟,知恵を理解したい.群れに秘められたからくりを理解したい.
過去の成果とそれに基づいて新たに湧き出したこのような問いに答えるため,当研究室では,新学術領域研究で「神経前駆細胞の動と静を制御する場と集団の原理」を課題として掲げている.「動」は,核移動であり,行き違い・すれ違いであり,分裂であり,押し・押されであり,また細胞産生モードの変化である.「静」は,apical面を維持する安定であり,幹細胞を枯渇させない安定である.
「見る」ことにもとづいて,問いのオリジナルさ・正当さを大切にし,現在を生きる研究者としてのベストを尽くしたい.
宮田卓樹 (2012年4月8日作成,4月16日更新)
(緑色論文は,当ラボまたは個別ページの「業績リスト」に一覧,オレンジ色の発表に関する情報も学会発表ページに一覧)
